Journal: Philosophie Jetzt – Menschenbild, ISSN 2365-5062, 27.Sept. 2018
URL: cognitiveagent.org
Email: info@cognitiveagent.org
Autor: Gerd Doeben-Henisch
Email: gerd@doeben-henisch.de
Gerald M.Edelman, Bright Air, Brilliant Fire. On the Matter of the Mind, New York: 1992, Basic Books
BISHER
Für die bisherige Diskussion siehe die kumulierte Zusammenfassung HIER.
KURZFASSUNG
Mit der Skizzierung seines Modells einer neuronalen Maschinerie liefert Edelman interessante Ansatzpunkte, um in einer möglichen, noch nicht existierenden, ‚Brückentheorie‘ zwischen Psychologie und Physiologie verschiedene psychologisch definierte Eigenschaften ‚von der Maschinerie her‘ plausibel zu machen. Eher unplausibel wirkt sein Versuch, die Wertefrage in Details des Genotyps und dann in Details des Phänotyps zu verlagern. Eine alternative Sicht wird vom Autor dieses Textes angedeutet.
KAP.9 NEURONALER DARWINISMUS
-
In diesem Kapitel will Edelman Argumente vorbringen, warum die ‚Gehirnwissenschaft‘ eine ‚Wissenschaft des Erkennens‘ bzw. ‚… der Erkennung‘ (‚recognition‘) ist.
-
Für seine Argumentation setzt Edelman voraus, dass wir Tiere (‚animals‘) haben, die sich in einer Umgebung befinden und sich in Interaktion mit dieser langfristig ‚anpassen‘, d.h. in der Lage sind, hinreichend viele Nachkommen überlebensfähig zu erzeugen. Diese Anpassungsfähigkeit beruht NICHT auf irgendwelche Instruktionen, sondern wird von den Tieren aufgrund der ihnen innewohnenden Eigenschaften ermöglicht. Diese innewohnende Eigenschaft, sich kurzfristig im Verhalten und langfristig in der Struktur des Körpers ‚anpassen zu können‘, verbindet Edelman mit dem Begriff ‚Erkennung‘ (‚recognition‘), der gebunden ist an die Struktur eines ‚Gehirns‘. Die von Edelman gemeinte Form der Erkennung basiert nicht auf expliziten Instruktionen sondern auf ‚Selektion/ Auswahl‘, die sich im Rahmen des Verhaltens ergibt. Dieses Verhalten muss in Verbindung mit dem Gehirn von der Gehirnwissenschaft untersucht werden. (vgl. 81f)
- In der Umgebung, die Edelman für seine Tiere voraussetzt, gibt es Ereignisse, die ’neu‘ sind, ‚unvorhergesehen‘! Solche Ereignisse kann man nicht mit schon bekanntem Wissen oder bekannten Regeln bearbeiten. Edelman postuliert daher, dass ein System, das auf ‚Auswahl/ Selektion‘ basieren soll, in der Lage ist, vor neuen Ereignissen in der Umgebung intern, im System, ‚Varianten‘ bilden kann. Die bisherigen Forschungen zur embryonalen und anschließenden Entwicklung des Gehirns legen nahe, dass dieser Prozess der ‚Gehirnentstehung/ Gehirnwerdung‘ keine 1-zu-1 Kopie eines vorgegebenen Plans ist, sondern ein Realisierungsgeschehen umfasst, bei dem sich in einer Mischung aus Umgebungsreizen und Zufällen neue individuelle Strukturen ausbilden, so dass selbst eineiige Zwillinge über unterschiedliche Feinstrukturen verfügen. Damit liegen unterschiedliche Gehirne vor, die aufgrund ihres im Körper umgesetzten Verhaltens selektiert werden können. Solch eine Variabilität wird darüber hinaus nicht nur einmal, zu Beginn des Lebens eines Tieres benötigt, sondern während des ganzen Lebens. Wie kann ein Gehirn dies leisten? (vgl. S.82)
- Innerhalb der Verhaltensanforderungen benennt Edelman einige Verhaltenseigenschaften explizit, die das Gehirn zu leisten in der Lage sein muss: (i) ‚Kategorisierung‘ von Ereignissen in der Wahrnehmung — auch schon vor-sprachlich –, (ii) Kategorisierungen im Kontext des Gedächtnisses, (iii) Aufbau eines ‚Bewusstseins‘, (iv) Entwicklung von ‚Sprache‘. (vgl. S.82)
-
Edelman formuliert dann – quasi zwischen drin – ein wissenschaftsphilosophisches Postulat, dass eine Theorie, die wissenschaftlich sein will, ausschließlich auf Prozessen und Ordnungen basieren muss, die in der physikalischen Welt (‚physical world‘) vorkommen. Und aus diesem wissenschaftsphilosophischen Postulat leitet er dann ein anderes wissenschaftsphilosophisches Postulat ab, dass nämlich eine wissenschaftliche Erklärung von ‚psychologischen‘ Prozessen aufzeigen muss, wie diese mit ‚physiologischen‘ Prozessen in Zusammenhang stehen.(vgl. S.82)
-
Eine solche wissenschaftliche Theorie zur Erklärung des Zusammenhangs zwischen psychologischen und physiologischen Prozessen hat Edelman in der ‚Theorie der neuronalen Gruppen Selektion (TNGS)‘ formuliert. Diese Theorie wurde zudem speziell entwickelt, um das Problem des ‚Bewusstsein‘ (‚consciousness‘) in den Griff zu bekommen. (vgl. S.82f)
-
Die beiden zentralen theoretischen Konzepte in der TNGS sind ‚TNGS Wiedereintritt‘ (‚TNGS reentry‘) und ‚Globales Abbilden‘ (‚global mapping‘). (vgl. S.83)
-
Im Zusammenhang damit muss man die Evolution neuer Formen (‚morphologies‘) sowohl vom Körper als auch vom Gehirn berücksichtigen. (vgl. S.83)
- Solche neuen Formbildungen finden im wesentlichen in drei Phasen statt: (i) die Entstehung von Netzwerken von Neuronen während der Entwicklung; (ii) unterschiedliche Gewichtungen der Verbindungen unter dem Einfluss von Einwirkungen von außen (und innen); (iii) Bildung von neuronalen Karten, die untereinander durch Wiedereintrittsverbindungen verbunden sind. (vgl. S.83)
- Bezüglich der frühen Entstehung von Netzwerken von Neuronen während der Entwicklung (Pase 1) wiederholt er die Feststellungen von vorausgehenden Kapiteln. Die Kernbotschaft ist, dass die sich herausbildenden Netzwerke genetisch nur sehr grob vor-bestimmt sind. Jedes einzelne Gehirn ist in dieser Phase sehr individuell. Die in dieser Phase verfügbaren Funktionen des Gehirns bilden ein ‚erstes Repertoire‚. (vgl. S.83)
- Mit der beginnenden Interaktion der Neuronen untereinander und durch Ereignisse an den Rändern (sensorische Wahrnehmung, propriozeptive Ereignisse im Körper,…) (Phase 2) werden einige Verbindungen ‚gestärkt‘, andere bleiben schwach, und mit diesen ‚Stärkungen von Verbindungen‘ bilden sich ’neue Funktionen‘ heraus, die dann insgesamt das ‚zweite Repertoire‚ bilden. (Vgl. S.83-85)
-
Dadurch dass neuronale Karten reziprok miteinander verknüpft sind (Phase 3) können die gleichen Ausgangsreize nicht nur parallel auf unterschiedliche Weise bearbeitet werden, sondern es ist auch möglich, dass sie sich im gleichen Zeitfenster wechselseitig beeinflussen und diese Signale in übergeordneten Karten zusammen geführt werden.(vgl. S.85)
- In Verbindung mit Gedächtnis (‚memory‘) sieht Edelman in diesen reziproken Karten nicht nur die Grundlage für Verhalten überhaupt sondern speziell auch für eine ‚Brücke‘ zwischen Physiologie und Psychologie. (vgl. S.85)
- An dieser Stelle führt Edelman nun – etwas überraschend – ‚neuronale Gruppen‚ (neuronal groups‘) ein als die eigentlichen Einheiten, die der ’neuronalen Selektion‘ unterliegen. Einzelne Neuronen eignen sich dafür schlecht, weil sie entweder nur ‚anregend‘ (‚excitatory‘) sind oder nur ‚hemmend‘ (‚inhibitory‘). Außerdem sendet ein einzelnes Neuron seine Synapsen zu vielen anderen Neuronen. Will man hier eine hinreichende Schärfe in der Adressierung realisieren, dann muss man viele Neuronen kombinieren, die sich in bestimmten Bereichen dann überlappen. Von daher können eigentlich nur ‚Gruppen von Neuronen‘ solche Anforderungen erfüllen und bieten sich von daher als natürliche Einheiten der Selektion an. (vgl. S.85-87)
-
Edelman will diese Themen dann am Beispiel der ‚Kategorisierung in der Wahrnehmung‘ (‚perceptual categorization) weiter ausführen. Er verweist dabei auch auf das Postskript mit einem Abschnitt zur Kategorisierung (S.233f). In diesem Postskript-Abschnitt kritisiert er vor allem jene Positionen, die bei der Frage der Kategorien von festen Strukturen ausgehen, die auch als solche ‚repräsentiert werden. Mit Verweis auf Wittgenstein und einigen Psychologen argumentiert er für eine Sicht der Kategorisierung, die sich aus der direkten Interaktion mit der physischen Umgebung ergibt. Ausgehend von konkreten Beispielen bilden alle Individuen konkrete Kerne, die sich im Laufe der Zeit erweitern, überlagern, ergänzen können. Einige der Elemente in solchen induktiven-dynamischen Netzen sind dann ‚typischer‘ als andere. Zwischen den Elementen können einige ‚ähnlicher‘ sein als andere, ’näher dran‘, usw. Aufgrund des induktiv-beispielhaften Charakters dieser Bildungsprozesse sind diese Strukturen bei allen Individuen grundsätzlich verschieden.
-
Das Phänomen der ‚Kategorisierung‘, das sich im Verhalten manifestiert (= Psychologie), gründet also in neuronalen Verschaltungen (= Physiologie), die im wesentlichen auf neuronale Karten (bestehend aus neuronalen Kernen) verweisen, die jeweils unabhängige Reize (Stimuli) empfangen, die aber aufgrund der reziproken Verbindungen sich innerhalb eines Zeitfensters wechselseitig bestärken. Mit anderen Worten, Eigenschaften A in der einen Karte werden zeitgleich mit Eigenschaften B in einer anderen Karte durch wechselseitige ‚Verstärkung‘ ’synchronisiert‘. Damit können verschiedene Eigenschaften zu einem Eigenschaftsbündel koordiniert werden. Je häufiger dies passiert, umso stärker wird diese Verbindung. Diese Synchronisierungsprozess können jederzeit mit beliebig vielen verschiedenen Karten passieren. Und, wie man erkennen kann, vollziehen sich diese Synchronisierungsprozesse nicht aufgrund eines ‚Supervisors höherer Ordnung‘, nicht aufgrund irgendwelcher ‚Instruktionen‘ von ‚außen‘, sondern aufgrund von physikalischen Außenweltereignissen (oder auch körperintern (propriozeptiv) erzeugte Ereignisse), die über Sensoren und neuronale Signalverarbeitung zu den Karten geleitet werden. Der Mechanismus der reziprok verschalteten Karten extrahiert aus diesen vor-verarbeiteten sensorischen (auch propriozeptiven) Signalen dann unterschiedliche Eigenschaften, die dann die ‚Bausteine für Kategorien‘ bilden. (vgl. S.87)
-
Da es ja sehr viele neuronale Karten mit sehr vielen reziproken Verbindungen untereinander gibt, können die Ausgänge von bestimmten Karten auch auf die Eingänge von vorausgehenden ‚Verarbeitungsstufen‘ zurück geleitet werden, um die Verarbeitung auf diese Weise zu modifizieren, oder diese reziproke Verbindungen werden für höhere Verarbeitungen benutzt, oder, oder , … Die Vielfalt der hier möglichen wechselseitigen Beeinflussungen ist groß. Edelman verweist auf ein Computermodell, das RCI (‚reentrant cortical integration‘) Modell (das reziproke kortikale Integrationsmodell), in dem er diese Theorie modellhaft umgesetzt und getestet hatte. Details dazu hat er in seinem Buch ‚The Remembert Past‘ beschrieben).(vgl. S.87-89)
-
Für die Kopplung des wahrgenommenen (perzeptuellen) Inputs eines Systems zu seinem Output (meist über Motorkomplexe), führt Edelman ferner noch ‚globale neuronale Karten‘ ein. Diese können den Output vieler lokaler neuronaler Karten zusammenfassen, integrieren, und damit komplexe Muster zur Steuerung von Motorkomplexen und damit dem beobachtbaren Verhalten nutzen. Zugleich können diese Outputs aber auch wieder rückgekoppelt werden auf den grundlegenden Input zum ‚(neuronalen) Vergleich‘, ob tatsächlich die ‚Wirkung‘ entsteht‘, die aufgrund des Outputs ‚(neuronal) erwartet‘ wird. Auf diese Weise kann die Gehirnwissenschaft (= Physiologie) eine neuronale Maschinerie anbieten, mit denen die Psychologie das äußerlich beobachtbare Verhalten ‚erklären‘ können.(vgl. S.87-89)
-
An dieser Stelle führt Edelman den Aspekt des ‚angemessenen‘ Verhaltens ein, das aus Sicht der Evolution und ‚Fitness‘ ‚besser geeignet ist‘ als ein anderes. Da er den Aspekt des ‚Supervisors‘ zuvor schon ausgeschlossen hat und auch keine ‚vorgegebenen Instruktionen‘ zulassen will, ist hier nicht ganz klar, wie in diesen hochdynamische induktive Struktur genetische ‚Präferenzen für Verhalten‘ eingeführt werden können. Er postuliert hier ‚interne Kriterien (‚internal criteria of value‘) dafür, dass Verhaltensweisen ‚werthaft‘ sind. Diese internen, genetisch bedingten, Kriterien bestimmen nicht konkrete Werte, sondern sie ‚beschränken die Bereiche‘ (‚constrain the domains‘), in denen Kategorisierungen stattfinden. Und er sieht diese genetisch bedingten Beschränkungen besonders in jenen Bereichen des Gehirns verortet, wo lebenswichtige Funktionen reguliert werden. Explizit nennt er: Herzschlag, Atmung, sexuelle Reaktion, Ernährungsreaktion, Drüsenfunktionen (= Ausschüttung chemischer Signalstoffe zur Steuerung von Funktionen), autonome (‚autonomic‘) Reaktionen. Diese stark vor-geprägten Kategorisierungen soll sich in einem entsprechend lebensförderndem Verhalten manifestieren.(vgl. S.90f)
-
Um diesen noch recht abstrakten Gedanken von ‚genetisch motivierter Präferenz‘ zu präzisieren berichtet Edelman von einem Computerexperiment ‚Darwin III‘, in dem im Rahmen des visuellen Systems solche neuronale Gruppen/ Karten ‚bevorzugt‘ wurden, die Licht verarbeiteten, das auf zentrale Bereiche des Auges traf. (vgl. S.91-93)
-
Unklar bleibt hier, wie diese ‚Bevorzugung‘ (‚Präferenz‘) überhaupt in das System kommt, so, wie es bislang von Edelman geschildert wurde. In der weiteren Erklärung zeigt Edelman einen größeren Systemzusammenhang von einem Auge, einem Arm, und das ‚Erlernen‘, einem visuellen Reiz einer bestimmten Art zu folgen und es zu greifen. Hier wird deutlich, dass sehr viele Subsysteme sich miteinander koordinieren müssen, damit Kategorisierungsleistung und Bewegung ‚zusammen passen‘. Es gibt auch eine erste Idee, wie sich diese Subsysteme über reziproke Verbindungen wechselseitig induktiv beeinflussen können. Wie ein zusätzlicher genetisch motivierter Mechanismus der ‚Bevorzugung‘ in dieses System Eingang findet, bleibt aber weiter unklar.(vgl. S.93)
-
Das ‚Präferenz (= Wert) System‘ (‚value system‘) ist klar zu unterscheiden von dem ‚Kategorisierung-System‘. Während das Kategorisierung-System induktiv über reziproke Karten arbeitet und darin ‚epigenetisch‘ wirksam ist, liegt das genetisch motivierte Präferenz-System dem epigenetischen Kategorisierung-System voraus. Edelman berichtet, dass im Computerexperiment, wenn man das Präferenz-System ‚heraus nimmt‘, dass dann das System nicht zum Ziel kommt. Damit postuliert er ein Axiom von der Notwendigkeit von Präferenz-Systemen für Kategorisierung-Systemen, ohne dass ganz klar wird, wie diese genetische motivierten Präferenzen in das System hineinkommen.(vgl. S.94)
-
In den folgenden Abschnitten diskutiert Edelman diverse Kritiken an seiner TNSG Theorie und kommt auf den Seiten 96f nochmals explizit auf das genetisch motivierte Präferenz-System zurück, während er sich gegen Kritik von Crick verteidigt. Grundsätzlich sieht Edelman im Rahmen der Evolution zwei selektive Systeme: (i) das ‚Evolutive‘, das aufgrund der Selektion von Individuen durch Passung/ Nicht-Passung zur Umgebung stattfindet, und (ii) das ’somatische‘, das sich primär durch Passung/ Nicht-Passung der neuronalen Gruppen zu den Reizanforderungen. Das somatische System ist aber nicht isoliert/ neutral gegenüber den genetischen Vorgaben, die als Präferenzen (‚values‘) die Selektion von geeigneten neuronalen Gruppen und Verschaltungen mit steuern. Der Ansatzpunkt für eine Einwirkung der genetischen Präferenzen auf das somatische System ohne Direktiven im Details sieht er über die wechselseitigen Wirkungen von ‚Strukturen‘, die als solche ‚genetisch motiviert‘ sein können. Wie diese ‚Motivierung bestimmter Strukturen‘ stattfinden kann, ohne den induktiven Prozesse zu stören, ist nicht ganz klar geworden.(vgl. SS.94-97)
DISKUSSION FORTSETZUNG: HIER KAP.9
-
Die Verwendung des Ausdrucks ‚Erkennung/ Erkennen‘ (‚recogniton‘) in Nr. 2 erscheint mir sehr gewagt, da die übliche Verwendungsweise des Ausdrucks ‚Erkennung‘ ein menschliches Subjekt voraussetzt, das im Rahmen seines Bewusstseins und dem zugehörigen – weitgehend unbewussten – ‚Kognitiven System‘ die wahrgenommenen – äußerliche (Sinnesorgane) wie innerliche (Körpersensoren) – Ereignisse ‚interpretiert‘ auf der Basis der ‚bisherigen Erfahrungen‘ und im Rahmen der spezifischen Prozeduren des ‚kognitiven Systems‘. Das von Edelman unterstellte ’selektive System‘ bezieht sich primär auf Strukturen, die sich ‚evolutiv‘ ausbilden oder aber ’somatisch‘ auf die Arbeitsweise des Gehirns, wie es sich im ‚laufenden Betrieb‘ organisiert. Das somatische System ist aber Teil des weitgehend ‚unbewussten kognitiven Systems‘, das in der alltäglichen Verwendung des Begriffs ‚Erkennung‘ ohne jegliche Bezugnahme auf die Details der Maschinerie einfach voraussetzt wird. Im Alltag liegt hier die Aufspaltung der Wirklichkeit in den bewussten Teil und den unbewussten vor, dessen wir uns – normalerweise – im alltäglichen ‚Vollzug‘ nicht bewusst sind (daher ja ‚unbewusst‘). Wenn Edelman also hier einen Ausdruck wie ‚Erkennung‘ aus dem ‚bewussten Verwendungszusammenhang‘ einfach so, ohne Kommentar, für einen ‚unbewussten Verwendungszusammenhang‘ benutzt, dann kann dies nachfolgend zu nachhaltigen Verwirrungen führen.
-
Der Punkt (3) von Edelman ist absolut fundamental. An ihm hängt alles andere.
-
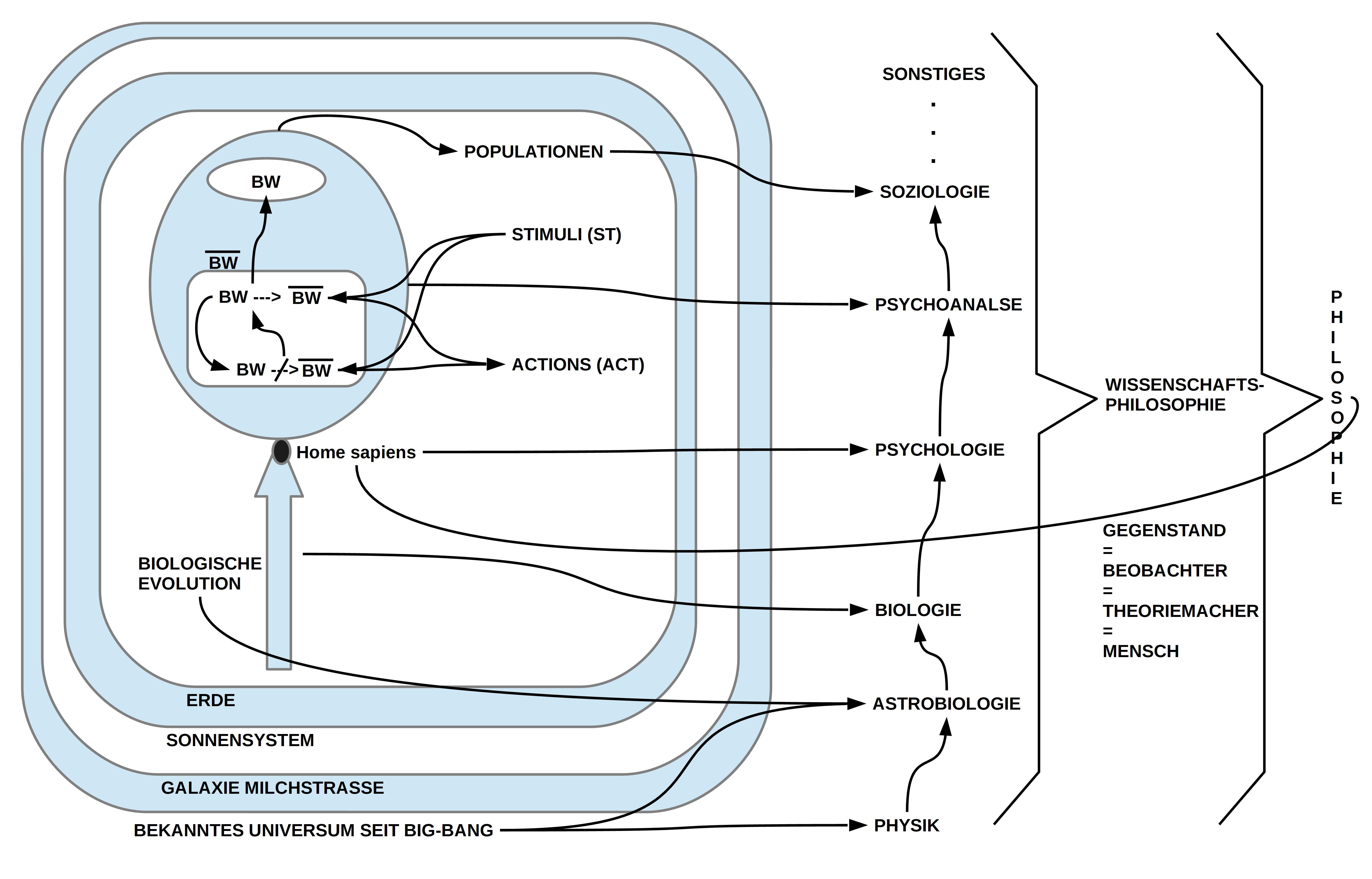
Der Punkt (4) spiegelt ein anderes fundamentales Verhältnis wieder, das Edelman immer und immer wieder thematisiert: das Verhältnis von Psychologie (die Wissenschaft vom Verhalten) und Physiologie (die Wissenschaft vom Körper, speziell vom Gehirn). Alle Eigenschaftszuschreibungen, die die Psychologie vornimmt (wie z.B. mit dem Ausdruck ‚Intelligenz‘, ‚Intelligenzquotient‘, ‚Reflex‘, usw.), basieren auf empirischen Beobachtungen von realem Verhalten von realen Menschen, die dann im Rahmen expliziter (formaler = mathematischer) Modelle/ Theorien in Beziehungen eingebracht werden. Die Psychologie schaut nicht (!) in das ‚Innere‘ des Systems. Das tut die Physiologie. Die Physiologie untersucht die Struktur und die Dynamik des Körpers; dazu gehört auch das Gehirn. Und auch die Physiologie sammelt empirische Daten von den Körperprozessen (z.B. wie ein Neuron zusammengesetzt ist, wie es physikalisch, chemisch, mikrobiologisch usw. funktioniert) und versucht diese empirische Daten in explizite Modelle/ Theorien einzubauen. Im Idealfall haben wir dann eine psychologische Theorie T_psych über beobachtbares Verhalten mit theoretisch unterstellten möglichen Faktoren ‚hinter‘ dem Verhalten, und parallel eine physiologische Theorie T_phys über körperinterne Prozesse, die die verschiedenen Einzelbeobachtungen in einen ‚Zusammenhang‘ bringen. Was wir aber NICHT haben, das ist eine Erklärung des Zusammenhangs ZWISCHEN beiden Theorien! Die Gegenstandsbereiche ‚beobachtbares Verhalten‘ und ‚beobachtbare Körperprozesse‘ sind zunächst voneinander völlig abgeschottet. Spontan vermutet man natürlich, dass es zwischen Gehirnprozessen, hormonellen Prozessen, Atmung usw. und dem beobachtbaren Verhalten einen Zusammenhang gibt, aber diesen Zusammenhang EXPLIZIT zu machen, das ist eine eigenständige Leistung mit einer ‚Brückentheorie‘ T_physpsych(T_phys, T_psych), in der dieser Zusammenhang ausdrücklich gemacht wird. Es gibt dazu sogar Ausdrücke wie ‚Psychosomatik‘, ‚Neuropsychologie‘, aber die dazu gehörigen Texte kommen bislang nicht als explizite ‚Brückentheorie‘ daher. Und wenn Edelman hier ausdrücklich die Ausdrücke ‚Wahrnehmung, ‚Gedächtnis‘, ‚Bewusstseins‘, ‚Sprache‘ nennt, dann handelt es sich ausnahmslos um hochkomplexe Sachverhalte, zu denen es bislang keine hinreichende psychologische Theorien gibt, wenngleich beeindruckende Forschungen und Forschungsansätze. Festzustellen, dass eine physiologische Theorie des Gehirns Erklärungsansätze liefern müsse, wie diese psychologischen Phänomene zu ‚erklären‘ sein ist folgerichtig, aber aktuell schwierig, da die Psychologie diese Phänomene noch keineswegs zweifelsfrei abklären konnte. Am weitesten fortgeschritten sich möglicherweise die psychologischen Forschungen zur ‚Wahrnehmung‘, zum ‚Gedächtnis‘, und irgendwie zu ‚Sprache‘, kaum zu ‚Bewusstsein‘ und dem Verhältnis zum ‚Unbewussten‘.
-
Die beiden in (5) aufgeführten metatheoretischen (= wissenschaftsphilosophischen) Postulate sind so, wie sie Edelman formuliert, mindestens missverständlich und darum Quelle möglicher gravierender Fehler. Die Verortung von wissenschaftlichen Aussagen in der ‚physikalischen‘ Welt und von speziell psychologischen Aussagen in den ‚physiologischen Prozessen des Körpers‘ sind als solche sicher kompatibel mit dem bisherigen Verständnis von empirischer Wissenschaft generell. Allerdings, wie gerade im vorausgehenden Punkt (C) dargelegt, gibt es empirische Datenbereiche (hier: Verhaltensdaten, Körperprozesse), die direkt zunächst nichts miteinander zu tun haben. Die auf diesen unterschiedlichen Datenbereichen beruhenden Modell-/ Theoriebildungen sind von daher zunächst UNABHÄNGIG voneinander und haben ihre wissenschaftliche Daseinsberechtigung. Zu wünschen und zu fordern, dass zwei zunächst methodisch unabhängige Modelle/ Theorien T_psych und T_phys miteinander in Beziehung gesetzt werden, das liegt ‚in der Natur des wissenschaftlichen Verstehenwollens‘, aber dies wäre eine zusätzliche Theoriedimension, die die beiden anderen Teiltheorien VORAUSSETZT. Dieser methodisch-logische Zusammenhang wird in der Formulierung von Edelmann nicht sichtbar und kann dazu führen, dass man eine wichtige Voraussetzung der gewünschten Erklärungsleistung übersieht, übergeht, und damit das Ziel schon im Ansatz verfehlt.
-
Dass – wie in (6) behauptet – die ‚Theorie der neuronalen Gruppen Selektion (TNGS)‘ jene Erklärung liefere, die die psychologischen Theorien ‚erklärt‘ ist von daher methodisch auf jeden Fall falsch. Allerdings kann man die Position von Edelman dahingehend verstehen, dass die TNGS ein physiologisches Modell liefere (was die große Schwäche besitzt, dass weder der übrige Körper, von dem das Gehirn nur ein verschwindend kleiner Teil ist, noch die jeweilige Umgebung einbezogen sind), das im Rahmen einer T_physpsych-Brückentheorie entscheidende Beiträge für solche einen Zusammenhang liefern könnte. Letzteres scheint mir in der Tat nicht ganz ausgeschlossen.
-
In den Punkten (7) – (12) beschreibt Edelman drei Phasen der Strukturbildung, wie es zu jenen physiologischen Strukturen kommen kann, die dann – unterstellt – dem beobachtbaren Verhalten ‚zugrunde‘ liegen. Allerdings, für das aktuell beobachtbare Verhalten spielen die verschiedenen Entstehungsphasen keine Rolle, sondern nur der ‚Endzustand‘ der Struktur, die dann das aktuelle Verhalten – unterstellt – ‚ermöglicht. Der Endzustand ist das Gehirn mit seinen neuronalen Verschaltungen, seinen Versorgungsstrukturen und seiner anhaltenden ‚Plastizität‘, die auf Ereignismengen reagiert. Aus den Punkten (7) – (12) geht nicht hervor, wo und wie diese Strukturen das ‚Bewusstsein erklären‘ können.
-
Wenn Edelman in (13) meint, dass seine neuronalen Karten mit den reziproken Verbindungen eine Grundlage für das psychologische Konzept des Gedächtnisses liefern können, dann ist dies sicher eine interessante Arbeitshypothese für eine mögliche Brückentheorie, aber die müsste man erst einmal formulieren, was wiederum eine psychologische Theorie des Gedächtnisses voraussetzt; eine solche gibt es bislang aber nicht.
-
Der Punkt (14) adressiert mögliche Details seiner Modellvorstellung von neuronalen Karten mit reziproken Verbindungen.
-
Der Punkt (15) greift das Thema von Punkt (13) nochmals auf. Die ‚Kategorisierung von Wahrnehmungsereignissen‘ ist letztlich nicht von dem Thema ‚Gedächtnis isolierbar. Er zitiert im Postskript auch viele namhafte PsychologenInnen, die dazu geforscht haben. Die Kerneinsicht dieser empirischen Forschungen geht dahin, dass sich solche Strukturen, die wir ‚Kategorien‘ nennen, ‚dynamisch‘ herausbilden, getriggert von konkreten Wahrnehmungsereignissen, die bei jedem individuell verschieden sind. Natürlich liegt hier die Vermutung nahe, dass entsprechend dynamische Strukturen im Körper, im Gehirn, vorhanden sein müssen, die dies ermöglichen. Dass hier Psychologie und Physiologie Hand-in-Hand arbeiten können, liegt auch auf der Hand. Aber dies stellt erhebliche methodische Herausforderungen, die Edelman kaum bis gar nicht adressiert.
-
In Punkt (16 – 18) beschreibt Edelman auf informelle Weise, wie er sich das Zusammenspiel von psychologischen Eigenschaften (die eine explizite psychologische Theorie T_psych voraussetzen) und seinem neuronalen Modell (für das er eine physiologische Theorie T_phys beansprucht) vorstellt. Auf dieser informellen Ebene wirkt seine Argumentation zunächst mal ‚plausibel‘.
-
Ab Punkt (19) berührt Edelman jetzt einen weiteren Fundamentalpunkt, der bis heute generell ungeklärt erscheint. Es geht um die Frage der ‚Werte‘, die ein Verhalten in einem gegeben Möglichkeitsraum in eine bestimmte ‚Richtung‘ anregen/ drängen …. (es gibt dafür eigentlich kein richtiges Wort!), so dass das System einem neuen Zustandsraum erreicht, der für es ‚günstiger‘ ist als ein anderer, auf jeden Fall ‚das Leben ermöglicht‘. Die Evolutionstheorie hat über das Phänomen der hinreichenden ‚Nachkommen‘ ein ‚operationales Kriterium‘ gefunden, mit dem sich der Ausdruck ‚Fitness‘ mit Bedeutung aufladen lässt. Über das ‚Überleben‘ bzw. ‚Nicht-Überleben‘ wirkt sich dies indirekt auf den Genotyp aus: die ‚günstigen‘ Genotypen überleben und die ’nicht günstigen‘ sterben aus. Ein ‚überlebender‘ Genotyp – einen, den man ‚fit‘ nennt – triggert dann den embryonalen Prozess und den darin gründenden ‚Entwicklungsprozess‘, der in einer ’nicht-deterministischen‘ Weise Körper- und damit Gehirnstrukturen entstehen lässt, die auf individueller Ebene immer verschieden sind. Jene Gehirnstrukturen, die für lebenserhaltende Systeme wichtig sind (letztlich die gesamte Körperarchitektur und das Zusammenspiel aller Komponenten untereinander) werden aber – trotz des nicht-deterministischen Charakters – offensichtlich dennoch herausgebildet. In all diesem lässt sich kein ‚geheimnisvoller Faktor‘ erkennen und er erscheint auch nicht notwendig. Das System ‚als Ganzes‘ repräsentiert ein ‚Erfolgsmodell‘. Hier jetzt – wie Edelman es versucht – im konkreten Details nach ‚Orten von Präferenzen‘ zu suchen erscheint unplausibel und seine Argumentation ist auch nicht schlüssig. Wenn Edelman ‚interne Kriterien für werthaftes Verhalten‘ postuliert, die genetisch bedingt sein sollen, dann postuliert er hier Eigenschaften, die sich nicht aus der Biochemie der Prozesse herauslesen lassen. Was er hier postuliert ist so eine Art von ‚genetischem Homunculus‘. Damit widerspricht er letztlich seinen eigenen wissenschaftlichen Überzeugungen. Letztlich ist es nur ein ‚Genotyp als ganzer‘, der ein ‚bislang erfolgreichen Phänotyp‘ repräsentiert. Scheiter er irgendwann – was auf lange Sicht der Normalfall ist, da die Erde sich ändert –, dann verschwindet der Genotyp einfach; jener, der ‚überlebt‘, verkörpert als ‚Ganzer‘ ein Erfolgsmodell, nicht ein einzelner, isolierter Aspekt. Mit dieser empirisch-wissenschaftlichen Sicht verschwinden die ‚Werte‘ zwar ‚aus dem System‘, aber das übergeordnete ’system-transzendente‘ Verhältnis zwischen System und Umwelt bleibt weiter erhalten und es ist dieses Verhältnis, das alleine darüber entscheidet, ob etwas ‚Leben-Ermöglichendes‘ ist oder nicht. Damit erweitert sich die Fragestellung nach der ‚Verortung möglicher Präferenzen‘ deutlich, wenn man das empirisch neue Phänomen des Bewusstseins einbezieht (das nur in Kombination mit einem Unbewussten auftritt!). Während die Umwelt-System-Beziehung primär über das Überleben entscheidet, ermöglicht das Bewusstsein in Kombination mit dem Unbewussten (z.B. dem Gedächtnis) eine ‚Extraktion‘ von Eigenschaften, Beziehungen, Dynamiken und mit kombinatorischen Varianten von dieser Umwelt-System-Beziehung, so dass eine kommunizierende Gruppe von Homo sapiens Exemplaren von nun an mögliche Zukünfte schon in der Gegenwart bis zu einem gewissen Graf ‚voraus anschauen‘ und auf ‚Lebensfähigkeit‘ ‚testen‘ kann. Insofern stellt das Bewusstsein und die Kommunikation eine radikale Erweiterung der Überlebensfähigkeit dar, nicht nur für den Homo sapiens selbst, sondern auch für die Gesamtheit des biologischen Lebens auf der Erde! Mit dem Ereignis Homo sapiens tritt das biologische Leben auf der Erde (und damit im ganzen Universum!?) in eine radikal neue Phase. Edelmans Versuch, ‚Werte‘ in einzelnen genetischen Strukturen und dann in einzelnen Strukturen des Gehirns zu finden wirken von daher unlogisch und abwegig. Edelmans Beispiele in den Punkten (20) – (23) können nicht klarmachen, wie diese interne Präferenzsystem funktionieren soll (ich werde diesen Punkt an konkreten Systemen nochmals selbst durchspielen).
-
Zusammenfassend würde ich für mich den Text von Edelman so interpretieren, dass (i) seine Skizze der neuronalen Maschinerie mit den reziproken Verbindungen und den unterschiedlichen Verschaltungen ganzer Karten ein interessanter Ansatzpunkt ist, für ein neuronales Modell, das in einer psychologisch-physiologischen Brückentheorie getestet werden könnte und sollte, sein Versuch aber (ii), die Wertefrage in biochemische Prozesse verorten zu wollen, ist in mehrfachem Sinne unplausibel, methodisch unklar und führt vom Problem eher weg.
Einen Überblick über alle Blogeinträge von Autor cagent nach Titeln findet sich HIER.
Einen Überblick über alle Themenbereiche des Blogs findet sich HIER.
Das aktuelle Publikationsinteresse des Blogs findet sich HIER.