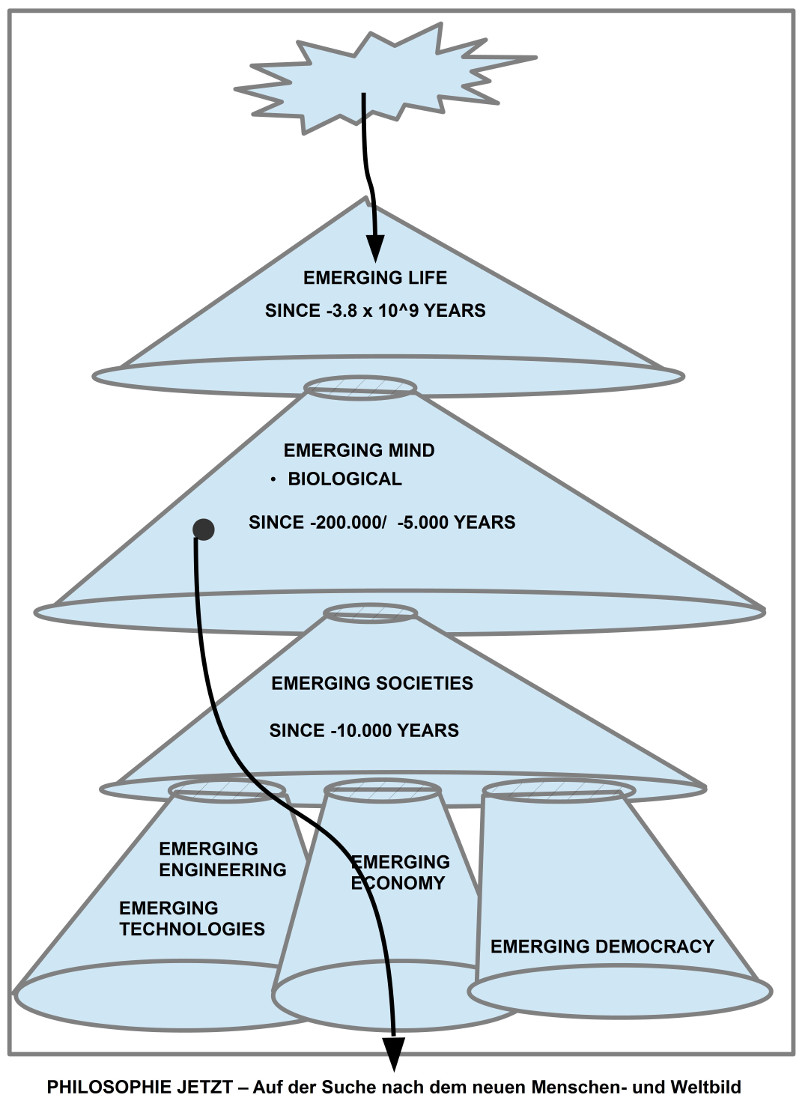
Journal: Philosophie Jetzt – Menschenbild, ISSN 2365-5062, 28.August 2018
URL: cognitiveagent.org
Email: info@cognitiveagent.org
Autor: Gerd Doeben-Henisch
Email: gerd@doeben-henisch.de
LETZTE ÄNDERUNG: 1.Dez.2018, Wiederholung der vorausgehenden Diskussion gestrichen. Kumulierte Zusammenfassungen sind HIER.
Gerald M.Edelman, Bright Air, Brilliant Fire. On the Matter of the Mind, New York: 1992, Basic Books
KAP.6 TOPOBIOLOGIE: WAS MAN VOM EMBRYO LERNEN KANN
- Der Ausgangspunkt ist weiterhin die Frage, welche materiellen Prozesse zu jenen materiellen Strukturen geführt haben, die für uns den homo sapiens repräsentieren. Diese materiellen Strukturen des homo sapiens zeigen eine Fülle von Verhaltenseigenschaften (Dynamiken), die wir als Hinweise auf ‚Geist‘ klassifizieren. Bisher wurden diese vorausgehenden formierenden Prozesse schon begrenzt auf die beiden Pole ‚Genotyp‘ und ‚Phänotyp‘ mit der Arbeitshypothese, dass die Eigenschaften des Genotyps weitgehend (wie weitgehend eigentlich? Kann man das quantifizieren?) die Eigenschaften des Phänotyps festlegen. Edelman selbst konkretisiert diese beiden Pole noch weitergehender mit der Frage, wie ein ‚ein-dimensionaler genetischer Kode‘ letztlich ein ‚drei-dimensionales Lebewesen‘ definieren kann.(vgl. S.63)
- Die Redeweise vom ‚genetischen Kode‘ setzt voraus, dass es eine Instanz gibt, die die Eigenschaften des Moleküls, das als ‚genetischer Kode‘ angesehen wird, als ‚Kode‘ ‚interpretieren‘ kann, d.h. diese Kode-erkennende-und-interpretierende Instanz (letztlich wieder ein Molekül) ist in der Lage, zwischen den materiellen Eigenschaften des Gen-Repräsentierenden Moleküls (normalerweise als DNA-Molekül vorliegend) und einer möglichen Proteinstruktur eine ‚Abbildung‘ vorzunehmen, die aus dem Bereich des ‚abstrakten Kodes‘ hinaustritt in den Bereich realer, 3-dimensionaler materieller Strukturen und Prozesse.
- Edelman beschreibt die konkreten Details des Gen-repräsentierenden-Moleküls M_gen (als DNA-Molekül; spezielle Abschnitte eines DNA-Moleküls repräsentieren ‚Gene‘), beschreibt die einzelnen Kode-Elemente (genannt ‚Kodons‘, ‚codons‘), die den späteren Transformationsprozess in materielle Strukturen steuern. Dieser Transformationsprozess geschieht aber nicht direkt, sondern über einen Zwischenschritt, in dem das Gen-repräsentierenden-Molekül M_gen in ein spiegel-identisches Gen-repräsentierendes-Molekül M_gen* (als RNA-Moleküle) übersetzt wird, das den Zellkern einer Zelle verlässt und dort dann von einer Kode-erkennende-und-interpretierende Instanz (‚cellular device‘) schrittweise in Aminosäuren übersetzt wird, die aneinander gekettet lange Moleküle (Polypeptide) bilden, die sich weiter als drei-dimensionale Strukturen ausformen, die schließlich Proteine repräsentieren. Aufgrund ihrer drei-dimensionalen Struktur kommen den Proteinen ‚Formen‘ (’shapes‘) zu, mit denen sich charakteristische ‚Eigenschaften‘, ‚Funktionen‘ verbinden. Schon diese Proteinformen kann man als Zwischenstufen zu einem Phänotyp ansehen. Unter anderem können sich Proteine zu komplexen ‚Zellen‘ zusammen finden (einem weiteren, komplexen Phänotyp), die ihren eigenen Zellkern haben mit einem spezifischen Genotyp. Verschiedene Proteine können ganz verschiedene Zellen bilden! (vgl. SS.52-57)
- Schon dieser Transformationsprozess von einem Gen-repräsentierenden-Molekül M_gen zu einer Zelle deutet in den einzelnen Phasen vielfältige Möglichkeiten der Variation, der Veränderung an. Doch, eine einzelne Zelle macht noch kein Lebewesen. Eine Lebensform wie der homo sapiens besteht – wie wir heute wissen – aus vielen Billionen (10^12) einzelnen Zellen, allein das Gehirn aus ca. 90 Milliarden neuronalen Zellen (ohne Glia-Zellen). Wie muss man sich diesen Weg von einer (!) befruchteten Zelle zu vielen Billionen Zellen in Form eines Lebewesens vorstellen?
- Dieser Transformationsprozess von einer befruchteten Eizelle ‚Zygote‘ genannt (‚zygote‘) zum ausgewachsenen Lebewesen wird von der Teilwissenschaft der ‚Embryologie‘ (‚embryology‘) behandelt. Die befruchtete Zelle (eine Vereinigung einer ‚Samenzelle‘ (’sperm cell‘) und einer ‚Eizelle‘ (‚egg cell‘) unterläuft eine lange Serie von Teilungen. Zellen können aber noch mehr als sich nur teilen: sie können ‚migrieren‘ (‚migrate‘), ‚absterben‘, ‚anhaften aneinander‘, und sich ‚differenzieren‘ (‚differentiate‘), d.h. abhängig von bestimmten chemischen Signalen in ihrer Umgebung werden unterschiedliche Gene aktiviert, um ganz spezifische Proteine zu erzeugen. Dadurch können Zellen (das Phänotyp) ganz unterschiedliche Dynamiken, unterschiedliche Verhaltensweisen zeigen. Differentiation von Zellen ist somit kontextabhängig, d.h. setzt die Nähe zu bestimmten anderen Molekülen (die als Signale fungieren) voraus, wobei die Signale Sender voraussetzen, die in Form von Zellen auftreten.(vgl. S.57f)
- Aufgrund dieser grundlegenden Fähigkeit der Differentiation (also eines signalabhängigen Verhaltens (Edelman spricht hier von ‚Ortsabhängigkeit‘; mit dem griechischen Wort ‚topos‘ für Ort spricht er dann auch von ‚Topobiologie‘; diese Wortwahl ist aber gefährlich, da sie primär Raumstrukturen nahelegt, obgleich der Prozess selbst nur Signale kennt (natürlich in einem Raum)) können Zellen, selbst wenn sie sich im Verlaufe des Transformationsprozesses zu unterschiedlichen ‚Schichten‘ (‚layer‘) anordnen, über Entfernungen hinweg ‚Signale senden‘, die kontextbedingt ausgelöst werden. Die schrittweise ‚Gestaltwerdung‘ von immer komplexeren Strukturen wird damit ‚aus sich selbst‘ gesteuert. Je nach aktueller Anordnung bestehen spezifische Kontexte, die spezifische neue Signale auslösen, die die ’nächsten Schritte‘ einleiten. Dabei spielen offensichtlich zwei Faktoren ineinander: (i) das in den Genen hinterlegte ‚Programm‘ konstituiert ein allgemeines ‚Schema‘, ein ‚Template‘, das einen grundlegenden Bauplan, eine grundlegende Architektur skizziert, und (ii) die jeweils konkreten individuellen Zellen, die assoziiert mit unterschiedlichen Kontexten durch den Raum wandern, bilden ein konkretes ‚Bedingungsgefüge‘, eine aktuelle ‚Selektionsmatrix‘, die darüber entscheidet, wann und wie Teile des generellen Bauplans konkretisiert werden. Der Möglichkeitsraum des genetischen Programms wird über die Konkretheit der selektierenden Zellen eingeschränkt, auf konkrete Punkte ‚herunter spezifiziert‘. Dieser Vorgang bietet zahlreiche Variationsmöglichkeiten. Mittlerweile hat man entdeckt, dass der Signalprozess stark hierarchisch sein kann insofern es ‚homöotische‘ (‚homeotic‘) Gene gibt, die die Proteinproduktion bestimmter Gene kontrollieren. (vgl. SS.58-63)
- Edelman benutzt den Begriff ‚Epigenetik‘ in den geschilderten Kontexten als jene Faktoren, Ereignisse, Prozesse, die zusätzlich zu den Genen selbst für die Umsetzung der Transformation von Genen in Proteinen, in Zellen, in Zellverbände verantwortlich sind.(vgl. S.62)
- Nach diesem embryologischen Einblick in die Entwicklungsdynamik eines Organismus kann Edelman skizzieren, wie die charakteristischen Strukturen des Gehirns eines homo sapiens sich gebildet haben können und warum diese Strukturen artspezifisch sind, d.h. diese Strukturen sind typisch für den homo sapiens; alle Exemplare der Lebensform ‚homo sapiens‘ haben die gleiche Struktur. Zugleich gilt aber auch, dass selbst eineiige Zwillinge sich im Detail dieser Strukturen unterscheiden können und tatsächlich unterscheiden. Das Zusammenspiel von allgemeinem genetischen Programm und den ‚konkreten individuellen Zellen‘ in ihren jeweils ’spezifischen Kontexten‘ bietet solch ungeheure Variationsmöglichkeiten, dass das gleiche artspezifische genetische Programm sich im Detail immer unterscheiden wird. Diese in der Art des Transformationsprozesses angelegte Variabilität ist aber eben nur die eine Seite. Die andere Seite ist die arttypische Struktur, die einen Organismus, ein Gehirn spezifiziert, wodurch eine ‚Architektur‘ definiert wird, die unterschiedlichste Funktionen an unterschiedliche ‚Komponenten‘ bindet, die zudem typische ‚Interaktionsmuster‘ zeigen. Exemplare des homo sapiens haben daher alle eine typische Weise des ‚Wahrnehmens‘, des ‚Erinnerns‘, des ‚Denkens‘, des ‚Fühlens‘, usw.(vgl. S.63f)
DISKUSSION FORTSETZUNG
- Die fundamentale Tatsache, dass es Moleküle gibt, die andere Moleküle als ‚Kode‘ benutzen können, um Transformationsprozesse zwischen einer Sorte von Molekülen (DNA, RNA) in eine andere Sorte von Molekülen (Polypeptide, Proteine) steuern zu können, nimmt Edelman als Faktum hin, thematisiert es selbst aber nicht weiter. Er benennt diese ‚interpretierenden Moleküle‘ auch nicht weiter; sein Begriff ‚cellular device‘ ist eher nichtssagend. Dennoch ist es gerade diese Fähigkeit des ‚Übersetzens’/ ‚Interpretierens‘, die fundamental ist für den ganzen Transformationsprozess von einem Genom in einen Phänotyp bzw. in eine ganze Kette von hierarchisch aufeinander aufbauenden Phänotypen. Setzt man diese Übersetzungsfähigkeit voraus, ist das ganze folgende Transformationsgeschehen – so komplex es im Detail erscheinen mag – irgendwie ‚trivial‘. Wenn ich in mathematischer Sicht irgendwelche Mengen habe (z.B. verschiedene Arten von Moleküle), habe aber keine Beziehungen definiert (Relationen, Funktionen), dann habe ich quasi ‚Nichts‘. Habe ich aber z.B. eine Funktion definiert, die eine ‚Abbildung‘ zwischen unterschiedlichen Mengen beschreibt, dann ist es eine reine Fleißaufgabe, die Abbildung durchzuführen (z.B. die Übersetzung von DNA über RNA in Aminosäuren, dann Polypeptide, dann Proteine). Das die Biochemie und Mikrobiologie samt Genetik so viele Jahre benötigt hat, die Details dieser Prozesse erkennen zu können, ändert nichts daran, dass diese Transformationsprozesse als solche ‚trivial‘ sind, wenn ich die grundlegende Transformationsfunktion definiert habe. Wo aber kommt diese grundlegende Abbildungsfunktion her? Wie kann es sein, dass ein ordinäres chemisches Molekül ein anderes ordinäres chemisches Molekül als ‚Kode‘ interpretiert, und zwar genau dann so, wie es geschieht? Betrachtet man ’normale‘ Moleküle mit ihren chemischen Eigenschaften isoliert, dann gibt es keinerlei Ansatzpunkt, um diese grundlegende Frage zu beantworten. Offensichtlich geht dies nur, wenn man alle Moleküle als eine Gesamtheit betrachtet, diese Gesamtheit zudem nicht im unbestimmten Raum, sondern in Verbindung mit den jeweils möglichen ‚realen Kontextbedingungen‘, und dann unter Berücksichtigung des potentiellen Interaktionsraumes all dieser Moleküle und Kontexte. Aber selbst diese Raum repräsentiert im mathematischen Sinne nur Mengen, die alles und nichts sein können. Dass man in diesem Raum eine Funktion implantieren sollte, die dem Dekodieren des genetischen Kodes entspricht, dafür gibt es im gesamten Raum keinerlei Ansatzpunkt, es sei denn, man setzt solche eine Funktion als ‚Eigenschaft des Raumes‘ voraus, so wie die Physiker die ‚Gravitation‘ als Eigenschaft des Raumes voraussetzen, ohne irgendeinen Ansatzpunkt im Raum selbst zu haben, als die beobachtbare Wirkung der Gravitation. Die Biologen können feststellen, dass es tatsächlich einen Transformationsprozess gibt, der solch eine Abbildungsbeziehung voraussetzt, sie haben aber keine Chance, das Auftreten dieser Abbildungsbeziehung aus den beobachtbaren materiellen Strukturen abzuleiten!!!
- In der Beschreibung von Edelmans Position habe ich schon angemerkt, dass seine Wortwahl ‚Topobiologie‘ möglicherweise unglücklich ist, da es letztlich nicht der dreidimensionale Raum als solcher ist, der entscheidend ist (wenngleich indirekt die Drei-Dimensionalität eine Rolle spielt) sondern der ‚Kontext in Form von interaktiven Nachbarschaften‘: welche andere Zellen stehen in Interaktion mit einer Zelle; welche Signale werden empfangen. Indirekt spielt dann auch der ‚vorausgehende Prozess‘ eine Rolle, durch den eben Kontexte erzeugt worden sind, die nur in bestimmten Phasen des Prozesses vorliegen. Man hat also eher einen ‚Phasenraum‘, eine Folge typischer Zustände, die auseinander hervorgehen, so, dass der bisherige Prozess die nächste Prozessphase hochgradig determiniert. Dies ähnelt einer ‚algorithmischen‘ Struktur, in der eine Folge von Anweisungen schrittweise abgearbeitet wird, wobei jeder Folgeschritt auf den Ergebnissen der vorausgehenden Abarbeitungen aufbaut und in Abhängigkeit von verfügbaren ‚Parameterwerten‘ den nächsten Schritt auswählt. Im Unterschied zu einem klassischen Computer, bei dem die Ausführungsumgebung (normalerweise) festliegt, haben wir es hier mit einem algorithmischen Programm zu tun, das die jeweilige Ausführungsumgebung simultan zur Ausführung ‚mit erschafft‘! Wenn Computermetapher, dann eben so: ein Programm (Algorithmus), das seine Ausführungsumgebung (die Hardware) mit jedem Schritt selbst ‚erschafft‘, ‚generiert‘, und damit seine Ausführungsmöglichkeiten schrittweise ausbaut, erweitert. Dies setzt allerdings voraus, dass das genetische Programm dies alles schon ‚vorsieht‘, ‚vorwegnimmt‘. Die interessante Frage ist dann hier, wie ist dies möglich? Wie kann sich ein genetisches Programm ‚aus dem Nichts‘ entwickeln, das all diese ungeheuer komplexen Informationen bezüglich Ausführung und Ausführungsumgebung zugleich ‚aufgesammelt‘, ’strukturiert‘, ‚verpackt‘ hat, wo die Gesamtheit der modernen Wissenschaft bislang nur Fragmente versteht?
- Während Neurowissenschaftler (Edelman eingeschlossen) oft mit unsinnigen Computervergleichen versuchen, die Besonderheit des menschlichen Gehirns herauszustellen, kann man ja auch mal umgekehrt denken: wenn die Entwicklung des Gehirns (und des gesamten Organismus) Ähnlichkeiten aufweist mit einem Algorithmus, der seine eigene Ausführungsumgebung während der Ausführung (!!!) mit generiert, ob sich solch ein Prozess auch ‚rein technisch‘ denken ließe in dem Sinne, dass wir Maschinen bauen, die aus einer ‚kleinen Anfangsmenge von Materie‘ heraus ausgestattet mit einem geeigneten ‚Kode‘ und einem geeigneten ‚Interpretierer‘ sich analog selbst sowohl materiell als auch kodemäßig entwickeln? Da die biologischen Systeme zeigen, dass es grundsätzlich geht, kann man solch einen technischen Prozess nicht grundsätzlich ausschließen. Ein solches Gedankenexperiment macht aber sichtbar, worauf es wirklich ankommt: eine solche sich selbst mit-bauende Maschine benötigt auch einen geeigneten Kode und Interpretationsmechanismus, eine grundlegende Funktion. Ohne diese Funktion, geht gar nichts. Die Herkunft dieser Funktion ist aber gerade diejenige grundlegende Frage, die die gesamte empirische Wissenschaft bislang nicht gelöst hat. Es gibt zwar neuere Arbeiten zur Entstehung von ersten Zellen aus Molekülen unter bestimmten realistischen Kontexten, aber auch diese Forschungen beschreiben nur Prozesse, die man ‚vorfindet‘, die sich ‚zeigen‘, nicht aber warum und wieso es überhaupt zu diesen Prozessen kommen kann. Alle beteiligten materiellen Faktoren in diesen Prozessen als solchen geben keinerlei Ansatzpunkte für eine Antwort. Das einzige, was wir bislang wissen, ist, dass es möglich ist, weil wir es ‚beobachten können‘. Die ‚empirischen Phänomene‘ sind immer noch die härteste Währung für Wahrheit.
Fortsetzung folgt HIER.
Einen Überblick über alle Blogeinträge von Autor cagent nach Titeln findet sich HIER.
Einen Überblick über alle Themenbereiche des Blogs findet sich HIER.
Das aktuelle Publikationsinteresse des Blogs findet sich HIER.